布鲁氏菌病(布病)是一种细菌性人畜共患传染病。世界大多数国家都有流行。由于带菌动物是其它动物和人类布病的主要传染源,因此,对动物的布病的防控就成为一个关键问题。
进入21世纪,布病在世界范围内呈现回升趋势。这一现象的出现固然与全球大流通时代的到来有密切关系,同时也表明防控技术和策略没能及时适应新的流行特点,没能达到理想的控制效果。
虽然,迄今有多种技术成熟,可用国际标准来标定的血清学方法可供诊断用。病原诊断也已经有成熟的分子生物学诊断技术。至少有4种技术成熟的布病疫苗可供选择。但在有效实施及技术本身的改进上还需进一步发展。
1布鲁氏菌病流行特点
1.1布病呈世界性流行
世界上大多数国家有过布病流行,1973年报道75个国家有疫情,1982年有94个国家,随着布病调查国家的增多,如今报道有布病发生的国家和地区达到160多个。经数十年努力,迄今在世界上已有14个国家和地区宣布清除了布病,有海峡群岛、挪威、瑞典、芬兰、丹麦、瑞士、捷克斯洛伐克、罗马尼亚、英国、荷兰、日本、奥地利、塞普路斯和保加利亚,另外,冰岛和维尔京群岛一直没有发现人畜布病存在。
自1990年以来,世界布病疫情在某些国家和地区也呈现了一定波动。如沙特阿拉伯于1987年发病人数为5283例,1989年上升为7893例,近几年已达年发病10000例。约旦于1985年仅有17例布病,1989年上升为628例,90年代后发病达1000例。1985年前土耳其布病年发病率为6/百万,1986~1989年间年发病率为30/百万,1990年后布病年发病率竟上升为90/百万。美国等发达国家的布病疫情也有一定波动。
羊布病为主要疫区的国家地区主要有地中海地区、非洲、东欧、南欧、西亚、中亚、东北亚、拉丁美洲。牛布病为主要疫区的国家地区主要有东欧、南欧、南美、亚洲。犬布病在美洲、亚洲、欧洲也呈散发存在,90年代欧洲还发现了海洋哺乳动物布病。
1.2布病呈再度流行状态
当中国布病疫情与世界布病疫情一样,呈再度肆虐状态。据1952~1990年的资料统计,中国人间布病疫情出现两个高峰(1957~1963年,1969~1971年),其发病率超过10/百万。70年代后至90年代初,人间布病发病率出现了明显下降,发病率降到了0.8/百万。自2000年后,我国人畜布病疫情出现了快速上升的势头。1993年全国新发病人数只有329例,迄今,全国布病疫情回升的省区已超过10个,2009年全国新发病例报告总数为35816例,报告发病率为2.70/10万。需要注意的是,人间的统计数据也仅仅主要是就诊数据统计,估算实际数据应该是统计数据的至少2倍以上。
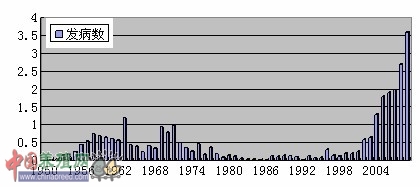
图1中国人间布病历史趋势
由于受经济转型、养殖规模的迅速扩大、兽医队伍建设不能满足需求等因素的影响,我国畜间布病疫情的统计数据不完整,但由于布病是一种以病畜传染人的人畜共患病,人间布病发生与家畜布病疫情是密切相关的。所以,上表人间病例的发展趋势,也是家畜疫情的发展趋势。
到2009年,除港澳台地区,我国家畜布病疫情按行政区域划分,基本呈华北、东北和西北严重流行,其余地区散发流行,仅海南无疫的状态。

图2全国人布病疫情扩散图
需要指出的是,上图人间布病病例严重的所在地并不表示是家畜布病的源头,只是表示此地的疫情比较严重。畜间病原可能来自外地,导致在当地家畜间传播,引起家畜疫情严重流行。
我国布病呈现明显的北高南低的,这与养殖品种和模式密切相关。北方是我国主要牧区,以羊牛成群放养为主,而羊牛是布病的主要宿主,放养比圈饲更易于病原传播。
布病的再度肆虐,很重要的一个原因是流通的飞速发展,造成传染源比以往任何时候都易于大范围传播。
1.3中国布病流行呈明显的季节特点
我国家畜布病呈现明显的春季高发的特点。这主要是因为我国北方以春季繁殖为主,而孕畜流产是布病的主要传播途径,一般在家畜应布病流产后一月左右的时间,出现感染家畜和人间病例的明显上升。而往南方,随着气侯的变暖,家畜怀孕季节的不明显,布病发生的季节性也变得不明显。
1.4中国布病流行呈明显的年龄与性别特点
对家畜来说,成年羊牛被感染的较多,特别是怀孕母畜。而性未成熟的幼畜有较强的抵抗性,感染较少。老年感染母畜常常流产现象减少,那是因为一方面空怀率上升,另一方面存在感染第一胎流产多见,第二胎后由于感染免疫导致流产率降低,但病原的传染性依然存在。
对人来说,青壮年男性病人居多,尤其是40至50年龄段的。因为在羊牛养殖量大的农牧区,40至50年龄段的男性是主要劳力,是接触流产物最多的人群。
1.5中国布病流行与养殖模式和人职业相关
我国主要羊牛养殖区域内,养殖模式以放养为主。与以往流行牧区高于农区不同,当前布病流行农区比牧区更重,并且呈现以农区向牧区扩散为主的特点。这种现象与农区家畜调进调出频繁有关,而牧区则以调出家畜为主,引进传染源的机率低于农区。
对人来说,当今的病例以养殖者为主,他们接触流产家畜的机率最高。其次是屠宰、贩运等与家畜及其畜产品频繁接触的人群。
1.6中国布病流行畜种有关
我国一直是羊种、牛种布鲁氏菌流行为主的国家。家畜中以羊布病最多发,其次是牛布病。羊种布鲁氏菌多发的主要原因在于羊只交易流动频繁,羊群中羊与羊接触频繁,另外羊种布鲁氏菌毒力和传染性最强也原因之一。牛也存在与羊类似的情况,因此我国牛布病疫情也相当严重。对人来说,羊种布鲁氏菌的毒力明显高于牛种菌,从人间分离到的病原菌90%以上是羊种布鲁氏菌,证实我国羊布病是人布病的主要源头。
当前,猪布病未像羊牛布病那样再度严重流行,是因为猪布病一直以自然配种为主要传播方式,感染对象主要是种猪。随着猪繁殖技术的发展,如今主要以人工授精方法繁殖,大大降低了猪间传染的机率。
2布病防控技术的发展
布病的防控技术主要包括2类,一类是检测技术,另一类是免疫技术。这2类技术自从19世纪发现布病病原以来,一直在不断发展。一些技术已相当成熟,在防控实践中得到广泛应用,并且帮助有些国家根除了该病。一些技术经过一段时期的评价后表明实用价值不大,逐渐被搁置。
2.1布病检测技术的发展
诊断技术一直是布鲁氏菌病研究的重要领域。在20世纪末的80、90年代,由于分子生物学的飞速发展,有力地推动了包括布鲁氏菌病在内的疫病诊断技术技术进步。
2.1.1病原学检测技术
至今,已发现布鲁氏菌有9个种。以牛种、羊种、猪种、犬种布鲁氏菌的流行最广,绵羊附睾型主要在澳洲流行,我国新疆曾经分离到,沙林鼠种只在美国局部存在,鲸种、鳍足种主要在欧美海洋动物中分离到。新近又分离到田鼠种布鲁氏菌。
表1布鲁氏菌8个种的常规分类鉴定特性表(GeoffreyFoster等2007年)

*参考菌株544和个别田间分离株为阴性。**少数菌株呈现裂解。
布鲁氏菌及其种与生物型的检测鉴定,除直接进行病原分离和上表的鉴定技术外,用过氧化物酶标记或荧光标记的特异性免疫染色检验技术已有报道。
90年代以来,分子生物学检测鉴定技术已得到大力发展。其中,PCR以其快速、准确、高效成为研究较多的检测方法。BetsyJBricKer和ShirleyMHalling在1994年针对牛、羊、绵羊、猪种布鲁氏菌设计不同的引物,应用PCR技术扩增在不同种间存在重复差异的布鲁氏菌保守重复基因单元IS711,以区别布鲁氏菌的种。1999年SerpeL等报道了用PCR一步法快速检测a软质干酪和牛乳中的布鲁氏菌。2000年SreevatsanS等报道用联合PCR对感染牛的乳汁、鼻分泌物检测布鲁氏菌和结核菌。ZygmuntMS等报道位于IS711序列下游的26bp基因是海洋哺乳动物的特殊标记。S19和RB51疫苗菌株已可以用PCR技术鉴定,S19的赤鲜醇敏感基因缺失702bp,所有其它布鲁氏菌都有1063bp,S19只有361bp。RB51菌株的编码葡胺聚糖酶的wboA基因发生了变异,在此基础上建立了RB51特异性PCR鉴定技术。PCR在布鲁氏菌的检测、鉴定方面越来越受到重视,以菌体基因组和基因组外的某些特定序列为检测基因,不仅可以确定布鲁氏菌的种,还可以确定属。此外,DNA探针技术、基因芯片技术等有望不久成为病原诊断的实用方法。
一些PCR技术已日趋成熟。AMOSPCR已被OIE确定为比较成熟的可以鉴别主要流行种的病原检测鉴定方法。RFLPPCR、脉冲凝胶电泳的技术]等已用于临床布鲁氏菌病原鉴定。
2.1.2免疫学检测技术
目前,我国防控实践中应用的比较成熟的免疫学检测技术主要有虎红平板凝集试验(RBT)、试管凝集试验(SAT)、补体结合试验(CFT)、全乳环状反应(MRT)。除SAT外,这些试验技术与国际上采用的技术是一致的。
传统方法的凝集试验(包括SAT和平板凝集试验)在发达国家已基本上停止使用,取代的方法是缓冲布鲁氏菌抗原试验(BBAT)、RBT。这两种试验除保持了快速凝集试验的高敏感性外,还由于采用低pH值而减少了非特异性反应。在国际贸易中,BBAT、RBT是牛、羊、猪种布鲁氏菌病诊断的指定试验,作为筛选试验用。
CFT至今仍是布鲁氏菌病的重要诊断方法,是牛、羊、绵羊副睾种布鲁氏菌病诊断的国际贸易指定试验,作为确诊试验用。也是公认的高特异性试验,但是由于试验操作繁杂,又难以标准化,并且敏感性降低,所以在实际防控中使用并不多。
MRT由于使用非常方便,并且具有良好的敏感性,是奶牛布鲁氏菌病监测的主要方法。
酶联免疫吸附试验(ELISA)是上世纪90年代发展起来的一种高敏感性的试验技术,而且操作更方便。ELISA既可以作为确定试验,又可以作为筛选试验。进入21世纪以来,OIE已将其列入国家贸易指定试验。ELISA方法不但用于血清学诊断,还可用于乳汁检查。猪种布鲁氏菌和绵羊副睾种布鲁氏菌亦已有成熟的ELISA诊断技术。关于ELISA的研究报道相当多,表明这是一种大家认为比较有前途的诊断方法。ELISA效果的关键在于抗原的选用。在已经标准化了的牛布鲁氏菌病ELISA诊断中,目前OIE推荐的是使用S-LPS抗原。有报道说,从牛种布鲁氏菌和羊种布鲁氏菌获得了能表达高度免疫原性蛋白的基因,其生产的蛋白可用于鉴别S19和Rev.1疫苗免疫与自然感染。用绵羊副睾种布鲁氏菌表面抗原及用重组蛋白和单克隆抗体作为ELISA抗原,认为可获得良好的诊断结果。对于ELISA技术来说,除所使用的抗原对技术的敏感性特异性有决定性影响外,还受到对敏感性特异性选择,当提高敏感性时,就会降低一定的特异性。
由于凝集试验不适合绵羊副睾种布鲁氏菌病的诊断,一种琼脂凝胶免疫扩散(AGID)试验就成为绵羊副睾种布鲁氏菌病的常用诊断方法。但未被采纳作国际贸易指定试验。
荧光偏振试验(FPA)是OIE认为敏感性、特异性与ELISA相当的试验,美国于2004年将FPA确定为官方确证试验。GallD等用荧光偏振法检测牛种布鲁氏菌,并与其它血清学检测方法进行了比较。认为荧光偏振法是检测牛布鲁氏菌最稳定的血清学方法,其敏感性和特异性在初步评价中分别为92.1%和99.4%,在后续的封闭试验中分别是96.3%和97.6%。通过双盲实验证实荧光偏振法还可以区别牛种布鲁氏菌感染和疫苗株S19免疫接种。MinasA等研究显示,FPA检测羊布病的敏感性和特异性分别可达到94.9%~97.7%和97.2%~98.4%。
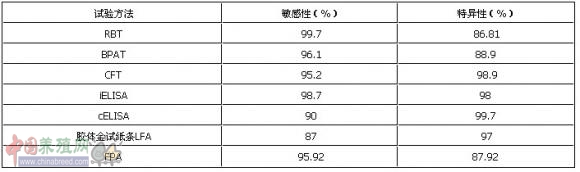
对于现用的主要布病血清学检测技术,不同的研究报道有着不同的结果,甚至有不同的结论。这与技术方法、样本数量、个体差异等因素有关。下表总结了部分研究报道显示的布病血清学检测技术的敏感性和特异性,总体来说,敏感性以RBT、iELSA、FPA为高,特异性以cELISA、CFT、iELSA为高。
表2不同试验方法的敏感性和特异性
2.2疫苗
目前世界范围内广泛应用的布鲁氏菌病疫苗有2种,S19和Rev.1。有些国家在用自己研制的疫苗,如中国的S2、M5活疫苗;俄罗斯的牛种布鲁氏菌82、82pn和75/79活疫苗。在美国阿拉斯加州的驯鹿,流行猪种布鲁氏菌生物型4。当地采用猪种布鲁氏菌生物型4佐剂灭活苗作为免疫预防用。
有些布鲁氏菌病疫苗曾经在较大范围内使用过,但由于经过一定时间的实际应用后,发现效果不理想,渐渐地就停用了或少用了,这样的疫苗有H38、45/20佐剂疫苗。
还有些疫苗经过短时间的实际应用,如1950年Kotlyarova研制的牛种布鲁氏菌104M活疫苗,1960年Yuskovets.M.K等研制的用于绵羊免疫的牛种布鲁氏菌68活疫苗,1952年Chernrshava等研制的用于绵羊免疫的猪种布鲁氏菌B61活疫苗,1956年Maiboroda研制的用于牛和猪的U.L.E.V。
曾经有人研究过混合疫苗。Cedro等(1961年),曾经在阿根廷使用过用活的牛种布鲁氏菌Viejo菌株与灭活的猪种布鲁氏菌菌株制备的混合疫苗。
自20世纪50年代开始,粗糙型布鲁氏菌疫苗一直是布鲁氏菌病疫苗研究的热点之一。报道实验室试验有效的不少于20种,从早期Jones等研制的R6、R7灭活佐剂疫苗,到后来的45/20灭活佐剂疫苗。最后都应免疫效果差或不是完全的无凝集原性而停止使用。但最近,美国研制出了一种粗糙型牛种布鲁氏菌活疫苗RB51,据报道有相当于S19疫苗的免疫效力,同时不干扰常规血清学诊断,在美国已获得政府批准正式使用。
当今疫苗免疫的最大问题是免疫干扰诊断检测。因此,新疫苗研究的重点集中在解决免疫对检测的干扰。同时,从分子生物学出发,对各种分子组分的抗原性和免疫原性的研究也相当活跃。
M.M.Kahl-McDonagh的研究显示,asp24基因缺失株对小鼠的保护率与S19相当。manBA,virB2缺失菌株也具有一定的免疫力。MARIAL.BOSCHIROLI、胡森等对缺失了bp26基因的S19、M5菌株研究表明,具有与父本相似的免疫原性。R.Adone对缺失了rpoB的粗糙型菌株研究显示,能达到几乎与Rev.1相似的免疫力。A.M.Arenas-Gamboa等的研究显示,vjbR缺失S19菌株有更好的安全性并保持了一定的免疫原性。除此以外,许多研究者对Cu/Zn、L7/L12、Omp10、Omp16、Omp19、Omp25、Omp31、virB8等基因缺失进行了研究,进一步了解布鲁氏菌的特性与功能,为研究新疫苗奠定了基础。此外,对于构建保护性抗原修饰疫苗,DNA疫苗也有研究。







 网友评论:(只显示最新10条。评论内容只代表网友观点,与本站立场无关!)
网友评论:(只显示最新10条。评论内容只代表网友观点,与本站立场无关!)

















{kind=link}